复能易锦提供的全部人类和小鼠ORF克隆及相关慢病毒和AAV病毒,所提供的载体质粒均采用毛细管电泳sanger全长测序验证,并承诺编码的氨基酸序列与NCBI数据库匹配。
All human and mice ORF clones and related Lentivirus, AAV particles and recombinant proteins provided by iGeneBio are fully sequenced using Sanger sequencing by capillary electrophoresis and Amino Acid Sequences are guaranteed to be matched with NCBI database.
HDR 供体克隆载体和定制的 HDR 供体克隆构建
。可用于通过同源重组介导的 CRISPR 基因组编辑应用。
Cas9稳定细胞系
。预制 Cas9 稳定表达细胞系特别适用于sgRNA文库筛选和其他高通量 CRISPR-Cas9 应用。
Safe Harbor 基因敲入试剂盒及 ORF 敲入克隆
。使用我们的人类 AAVS1 和小鼠 ROSA26Safe Harbor 基因敲入试剂盒可以轻松实现几乎任何DNA片段的成功敲入。人类及小鼠各有超过20,000条经过序列验证的ORF可用于转入基因的供体克隆设计。
IndelCheck™ 插入缺失检测体系
。IndelCheck™ CRISPR/TALEN 插入缺失检测体系是一套设计用于验证基因组编辑工具功能或检测编辑阳性细胞的完整体系,包括靶点 PCR 试剂盒、T7 核酸内切酶 I 检测试剂盒以及靶点 PCR 克隆试剂盒。另外,靶位点特异性PCR引物亦可选择。
sgRNA 文库
。我们提供7款预制的、通路特异性的 sgRNA 文库,每款都有慢病毒颗粒、转染级质粒 DNA 以及菌种等三种形式可供选择。客户也可以自行定制满足实验需求的 sgRNA 文库。
转基因小鼠服务
。GeneCopeia 提供原核显微注射法生成的、CRISPR介导的基因组修饰小鼠。基因组修饰的类型包括组成型基因敲除、条件性基因敲除、组成型基因敲入、人源化及转基因等。
VividFISH™染色体FISH计数探针
。FISH是帮助您鉴定靶基因拷贝数的强大工具。在进行基因组编辑实验前使用VividFISH™探针来鉴定目的染色体拷贝数,可对后续筛选目的修饰阳性克隆有所帮助。
GeneCopoeia为客户感兴趣的基因提供sgRNA(single-guide RNA)设计与克隆定制服务。sgRNA克隆表达一个单链的融合sgRNA(由crRNA和tracrRNA融合而成)。当Cas9核酸酶存在时,sgRNA能识别靶标DNA序列并引导Cas9核酸酶剪切靶标位点,形成DNA双链断裂(DSBs),进而实现基因敲除、敲入及突变等基因组编辑。多个sgRNA克隆与一个Cas9克隆共转染可同时在基因组中编辑多个位点,使实验设计更高效、更灵活。
sgRNA载体类型
sgRNA
是否含有Cas9核酸酶基因
筛选标记/报告基因
价格(¥)
SG001
pCRISPR-SG01
1 或多个
无,Cas9克隆单独出售*
Hygromycin
SG002
pCRISPR-LvSG03
1 或多个
无,Cas9克隆单独出售*
Puromycin / mCherry
SG012
pCRISPR-CG02
有,载体含有CBh启动子启动表达的Cas9核酸酶基因
SG013
pCRISPR-CG04
有,载体含有CMV启动子启动表达的Cas9核酸酶基因
Neomycin / copGFP
SG014
pCRISPR-CG07
有,载体含有CBh启动子启动表达的Cas9核酸酶基因
copGFP
SG015
pCRISPR-CG08
有,载体含有CBh启动子启动表达的Cas9核酸酶基因
mCherry
pCRISPR-CG12
有,载体含有CMV启动子启动表达的Cas9核酸酶基因
Neomycin / mCherry
*配合单独表达Cas9 核酸酶 (Cat.No.
CP-C9NU-01
,
CP-LvC9NU-01
,
CP-LvC9NU-02
,
CP-LvC9NU-08
,
CP-LvC9NU-09
,
CP-LvC9NU-10
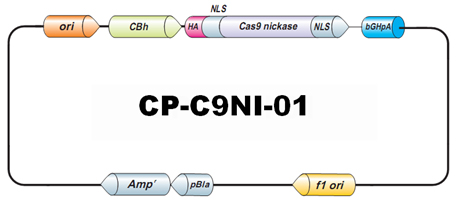
) 或 Cas9 D10A 切口酶 (Cat.No.
CP-C9NI-01
,
CP-C9NI-02
) 的 Cas9 克隆使用。
购买 货号 产品 筛选标记 报告基因 价格(¥)
CCPCTR01-CG02 Scrambled sgRNA control for pCRISPR-CG02. Scrambled sgRNA control for pCRISPR-CG02. N/A 1100
CCPCTR01-CG04 Scrambled sgRNA control for pCRISPR-CG04 Scrambled sgRNA control for pCRISPR-CG04 1100
CCPCTR01-CG07 Scrambled sgRNA control for pCRISPR-CG07 Scrambled sgRNA control for pCRISPR-CG07 1100
CCPCTR01-CG08 Scrambled sgRNA control for pCRISPR-CG08 Scrambled sgRNA control for pCRISPR-CG08 with mCherry. 1100
CCPCTR01-LvSG03 Scrambled sgRNA control for pCRISPR-LvSG03 Scrambled sgRNA control for pCRISPR-LvSG03 mCherry 1100
CCPCTR01-SG01 Scrambled sgRNA control for pCRISPR-SG01 Scrambled sgRNA control for pCRISPR-SG01 N/A 1100
LPP-CCPCTR01-LvSG03-100
sgRNA 对照 CCPCTR01-LvSG03 的纯化慢病毒颗粒(25 µL×4管)
10
8
TU/ml sgRNA 对照 CCPCTR01-LvSG03 的纯化慢病毒颗,转导级
Puromycin
CCPCTR01-LvSG03
¥1200
LPP-CCPCTR01-LvSG03-400
sgRNA 对照 CCPCTR01-LvSG03 的纯化慢病毒颗粒(100µL,25 µL×16管)
10
8
TU/ml sgRNA 对照 CCPCTR01-LvSG03 的纯化慢病毒颗,转导级
Puromycin
CCPCTR01-LvSG03
¥3200
图5. CRISPR-Cas9多重靶向编辑多个基因。
实验组(1-4泳道)为Cas9表达克隆及分别表达靶向p53, HUWE1, NCL3 和 GFP基因的sgRNA克隆质粒共转染HEK293t GFP稳转细胞。对照组(5-8泳道)为Cas9表达克隆质及随机sgRNA表达克隆的质粒共转染HEK293t GFP稳转细胞。通过T7核酸内切酶检测分析基因组DNA各个靶点上同时存在的插入缺失。*号标记的条带显示Cas9核酸酶及分别靶向多个靶点的sgRNA都在目标靶点上有效地诱发了插入缺失突变(1-4道)。PCR产物条带及T7核酸内切酶酶切产物条带大小:GFP: 720bp (完整), 340bp + 380bp (酶切); NCL3: 765bp (完整), 295bp +470bp (酶切); HUWE: 520bp (完整), 190bp + 330bp (酶切); P53: 825bp (完整), 475bp + 350bp (酶切).
Cas9表达克隆
GeneHero™ Cas9 核酸酶及 D10A 切口酶表达克隆
Cas9核酸酶克隆表达按照人类密码子优化过的Cas9核酸酶基因。Cas9切口酶克隆表达一个Cas9 D10A 切口酶基因。该切口酶是一个
Cas9突变体,第10位氨基酸从野生型的天冬氨酸突变成了丙氨酸。
CRISPR-Cas9体系是由sgRNA和Cas9核酸酶构成的DNA靶向识别剪切复合体。sgRNA识别一段20bp、3端紧接着5′-N-G-G-3′ PAM位点的DNA靶序列,Cas9核酸酶在该靶序列上生成DNA双链断裂(DSB)。DNA双链断裂(DSB)会被细胞的非同源末端连接(NHEJ)或同源重组(HR)机制修复。NHEJ机制容易在修复的断裂位点引入插入/缺失突变,造成阅读框移码而产生基因敲除的效果。同源重组(HR)需要细胞内同时存在一段带有靶点同源序列的供体DNA作为修复模板,我们可以通过对供体DNA进行设计,在众多实验体系中实现基因敲除或者敲入突变修饰、报告基因等等一系列基因组修饰。
Cas9 D10A切口酶跟野生型的Cas9一样,也使用sgRNA识别靶序列。但
Cas9 D10A切口酶在靶序列上生成的是单链切口而非双链断裂。单链切口通常会由NHEJ和HR以外的机制完美地修复。但通过设计一对定向正确的sgRNA分别识别靶点两侧的DNA双链,我们可以诱导Cas9 D10A分别在靶点两侧的DNA双链上各生成一个单链切口,构成一个双链断裂(DSB),进而诱导NHEJ或HR修复机制。这种双切口策略常被用在要求低脱靶率的实验设计中。
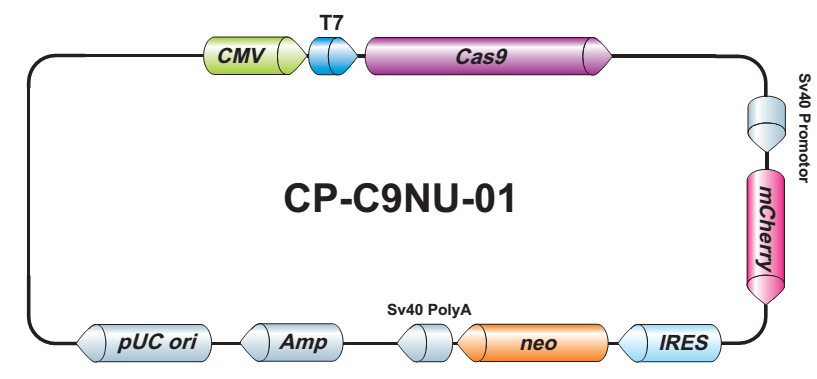
LPP-CP-LvC9NU-01-100
Cas9 核酸酶纯化 Lentifect™ 慢病毒颗粒
1×10^6 – 1×10^7 TU/ml Cas9 核酸酶慢病毒颗粒,转导级,100 µL.
Neomycin
CP-LvC9NU-01
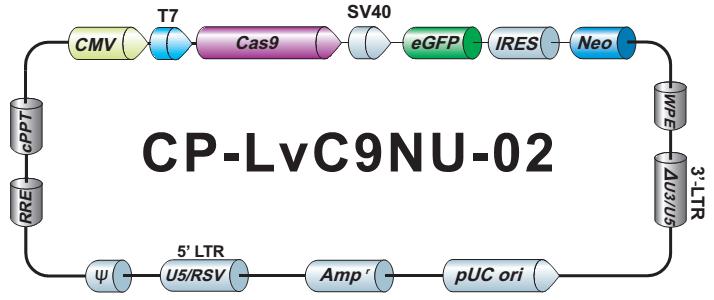
LPP-CP-LvC9NU-02-100
Cas9 核酸酶纯化 Lentifect™ 慢病毒颗粒
1×10^6 – 1×10^7 TU/ml Cas9 核酸酶慢病毒颗粒,转导级,100 µL.
eGFP/Neomycin
CP-LvC9NU-02
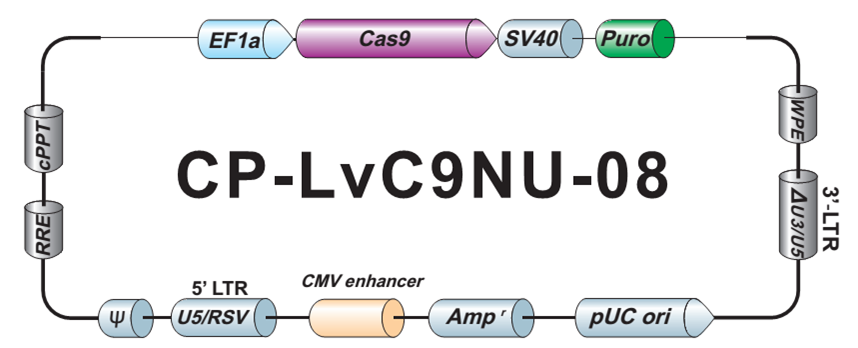
LPP-CP-LvC9NU-08-100
Cas9 核酸酶纯化 Lentifect™ 慢病毒颗粒
不低于1×10^7 TU/ml Cas9 核酸酶慢病毒颗粒,转导级,100 µL.
Puromycin
CP-LvC9NU-08
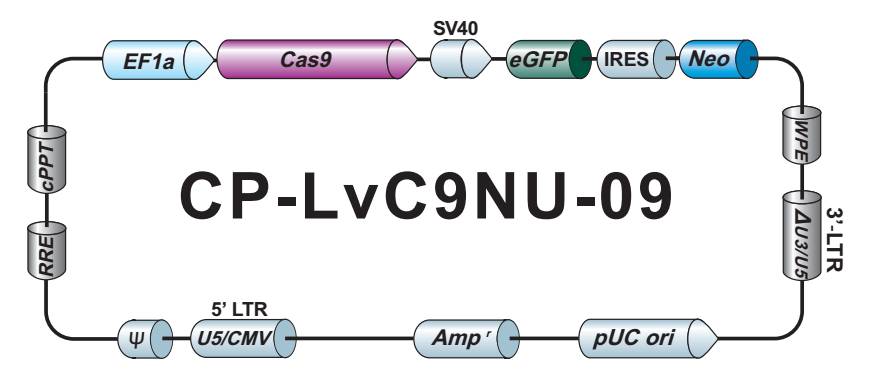
LPP-CP-LvC9NU-09-100
Cas9 核酸酶纯化 Lentifect™ 慢病毒颗粒
不低于1×10^7 TU/ml Cas9 核酸酶慢病毒颗粒,转导级,100 µL.
eGFP/Neomycin
CP-LvC9NU-09
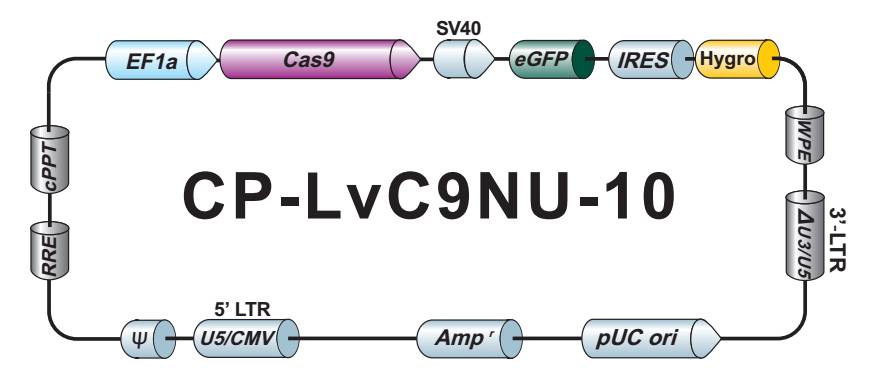
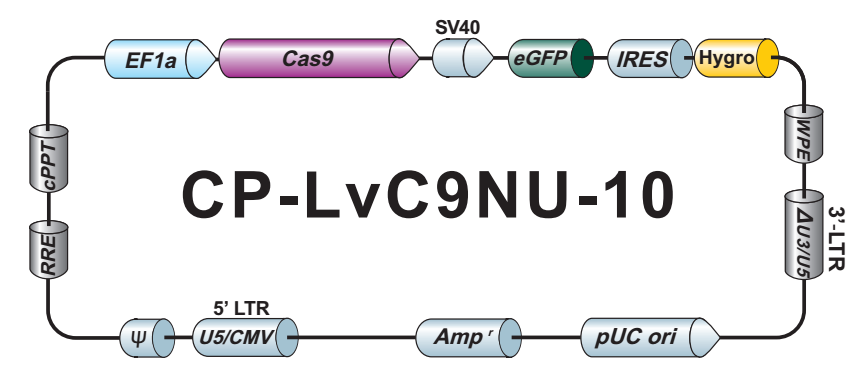
LPP-CP-LvC9NU-10-100
Cas9 核酸酶纯化 Lentifect™ 慢病毒颗粒
不低于1×10^7 TU/ml Cas9 核酸酶慢病毒颗粒,转导级,100 µL.
eGFP/Hygromycin
CP-LvC9NU-10
LPP-CP-LvC9NU-01-400
Cas9 核酸酶纯化 Lentifect™ 慢病毒颗粒
1×10^6 –1×10^7 TU/ml Cas9 核酸酶慢病毒颗粒,转导级,400 µL.
Neomycin
CP-LvC9NU-01
LPP-CP-LvC9NU-02-400
Cas9 核酸酶纯化 Lentifect™ 慢病毒颗粒
1×10^6 – 1×10^7 TU/ml Cas9 核酸酶慢病毒颗粒,转导级,400 µL.
eGFP/Neomycin
CP-LvC9NU-02
LPP-CP-LvC9NU-08-400
Cas9 核酸酶纯化 Lentifect™ 慢病毒颗粒
不低于1×10^7 TU/ml Cas9 核酸酶慢病毒颗粒,转导级,400 µL.
Puromycin
CP-LvC9NU-08
LPP-CP-LvC9NU-09-400
Cas9 核酸酶纯化 Lentifect™ 慢病毒颗粒
不低于1×10^7 TU/ml Cas9 核酸酶慢病毒颗粒,转导级,400 µL.
eGFP/Neomycin
CP-LvC9NU-09
LPP-CP-LvC9NU-10-400
Cas9 核酸酶纯化 Lentifect™ 慢病毒颗粒
不低于1×10^7 TU/ml Cas9 核酸酶慢病毒颗粒,转导级,400 µL.
eGFP/Hygromycin
CP-LvC9NU-10
GeneHero™ Cas9 稳定表达细胞系
GeneCopoeia 提供 Cas9 核酸酶的稳定表达细胞系。这些细胞系能为您的 CRISPR 基因组编辑应用提供一个方便、高效的工具。
我们会提供预制的GeneHero™ Cas9 人类或小鼠稳定表达细胞系,如H1299或HEK293T。Cas9 核酸酶的基因已被稳定整合到这些细胞系基因组上的 safe harbor 位点(图1)。此外,我们也能将 Cas9 核酸酶基因稳定整合到您所指定的细胞系基因组。
更多产品信息,请访问我们的
Cas9稳定表达细胞系
产品页面。
GeneCopoeia 也提供稳定细胞系构建服务。稳定细胞系构建服务请联系以下邮箱
inquiry@igenebio.com
,或填写以下
表格
。
CRISPR工作原理
CRISPR-Cas系统(The clustered, regularly interspaced, short palindromic repeats-associated protein systems)是一个细菌及古细菌进化出来用以抵御病毒和质粒入侵的适应性机制。CRISPR-Cas系统的高效基因组编辑功能已被应用于多种生物,包括斑马鱼、小鼠、大鼠、秀丽隐杆线虫、植物及细菌。多个科研小组的研究都显示,与锌指核酸酶(ZFNs)和转录激活样效应核酸酶(Transcription activator-like effector nucleases, TALEN)相比较,CRISPR-Cas系统介导的基因组靶向实验在细胞或斑马鱼中具有相似甚至更高的效率。
CRISPR-Cas9体系的RNA-DNA识别机制为基因组工程研究提供了一项简便而强大的工具。该体系其中一个最重要的优势是Cas9蛋白可在多个不同的gRNA的引导下同时靶向多个基因组位点。
图 1.
CRISPR/Cas9介导的基因组编辑原理图
在II型CRISPR系统中,CRISPR RNA(crRNA)与转录激活crRNA(Trans-activating crRNA, tracrRNA)退火形成的复合物能特异识别基因组序列,引导Cas9核酸内切酶在目的片段生成DNA双链断裂(double-strand breaks, DSBs)。 这个识别复合体可以通过融合crRNA与tracrRNA序列形成sgRNA(single-guided RNA)进行简化。基因组的靶序列中有长约20bp的片段与crRNA或sgRNA互补配对;靶序列末端的三核苷酸区域PAM(5’-NGG-3’)为Cas9识别位点,是实现剪切功能的关键。
图2. CRISPR-Cas9介导的基因编辑。
(左):sgRNA引导Cas9核酸酶作用于基因组,形成的DSBs被非同源末端连接(NHEJ)机制修复;(右):sgRNA引导Cas9核酸酶作用于基因组,形成DSBs,同源重组(HR)作用下,供体质粒上的目标基因及筛选标记(或其他遗传元件)在断裂处被整合进基因组。
图3:
切口酶在结合链生产单链切口的原理图
TALEN
与 CRISPR-Cas9 体系对比
Horvath P, Barrangou R (January 2010). “CRISPR/Cas, the immune system of bacteria and archaea”. Science 327 (5962): 167–70.
Marraffini LA, Sontheimer EJ (February 2010).
“CRISPR interference: RNA-directed adaptive immunity in bacteria and archaea”
.
Nat Rev Genet
11 (3): 181–190.
Hale CR, Zhao P, Olson S, et al. (November 2009). “RNA-Guided RNA Cleavage by a CRISPR RNA-Cas Protein Complex”. Cell 139 (5): 945–56.
van der Oost J, Brouns SJ (November 2009). “RNAi: prokaryotes get in on the act”. Cell 139 (5): 863–5. doi:10.1016/j.cell.2009.11.018.
Hale CR, Zhao P, Olson S, et al. (November 2009).
“RNA-Guided RNA Cleavage by a CRISPR RNA-Cas Protein Complex”
.
Cell
139 (5): 945–56.
Jinek, M., Chylinski, K., Fonfara, I., Hauer, M., Doudna, J.A., and Charpentie E. (2012). A programmable dual-RNA-guided DNA endonuclease in adaptiv bacterial immunity. Science 337, 816–821.
Jiang, W., Bikard, D., Cox, D., Zhang, F., and Marraffini, L.A. (2013). RNA-guided editing of bacterial genomes using CRISPR-Cas systems. Nat.Biotechnol. 31, 233–239.
Hsu, P.D., Scott, D.A.,Weinstein, J.A., Ran, F.A., Konermann, S., Agarwala, V.,Li, Y., Fine, E.J., Wu, X., Shalem, O., et al. (2013). DNA targeting specificity of RNA-guided Cas9 nucleases. Nat. Biotechnol. Published online July 21, 2013.
Ran, et al. (2013). Double Nicking by RNA-Guided CRISPR Cas9 for Enhanced Genome Editing Specificity. Cell 154, 1380.
GeneHero™ CRISPR-Cas9 用户手册
基因组编辑常见问题及CRISPR FAQ
Protocols
CRISPR Protocol 1: Gene Knockout Without Donor
CRISPR Protocol 2: HDR Donor Plasmid Applications (gene knockout, gene mutagenesis, gene tagging, Safe Harbor ORF knock-in)
Knockout By TALEN Or CRISPR vs. Knockdown By shRNA or siRNA
CRISPR-Cas9 Specificity: Taming Off-target Mutagenesis
Genome Editing: Which Should I Choose, TALEN Or CRISPR?
Genome Editing In Mammalian Cells: What Do I Do Next?
Genome Editing: HDR Donors For Gene Knockout, Mutagenesis, Tagging, and Safe Harbor Knock-in
Genome Editing: Applications For GeneCopoeia CRISPR sgRNA Libraries
IndelCheck
™
: A Powerful CRISPR/TALEN Validation & SCreening Tool
Genome Editing: Cas9 Stable Cell Lines for CRISPR sgRNA Validation, Library Screening, and More
Teng, Y., et al. (2017).
MVP-mediated exosomal sorting of miR-193a promotes colon cancer progression
. Nature Communications doi: 10.1038/ncomms14448 [
Lentiviral particles expressing sgRNA targeting MVP
]
Qi, C. et al. (2016)
HEK293T Cells Are Heterozygous for CCR5 Delta 32 Mutation
. PLOS ONE, April 4, 2016 http://dx.doi.org/10.1371/journal.pone.0152975. [
T7E1 nuclease
].
Hsu, PY. et al. (2016)
Spatiotemporal control of estrogen-responsive transcription in ERα-positive breast cancer cells
. Oncogene, 35, 2379–2389; doi:10.1038/onc.2015.298; published online 24 August 2015.
[DEREs, sgRNA contructs and validation
].
Blackburn, JB. Et al. (2016)
COG Complex complexities: Detailed characterization of a complete set of HEK293T cells lacking individual COG subunits
. Frontiers in Cell and Developmental Biology, 2016; 4: 23.
[Cog5, Cog2, Cog7 sgRNA].
Hu, B., et al. (2016).
A Novel Missense Mutation of CMT2P Alters Transcription Machinery
. Annals of Neurobiology doi: 10.1002/ana.24776
[LRSAM1 CRISPR sgRNA clone]
Bahal, R., et al. (2016).
In vivo correction of anaemia in β-thalassemic mice by γPNA-mediated gene editing with nanoparticle delivery.
Nature Communications doi: 10.1038/ncomms13304 [
Cas9 and sgRNA clones targeting mouse b-globin]
Zhang, J. et al. (2015)
Focal adhesion kinase-promoted tumor glucose metabolism is associated with a shift of mitochondrial respiration to glycolysis
. Oncogene, 35, 1926-1942 (14 April 2016) | doi:10.1038/onc.2015.256-. [
CRISPR-Cas9 genome editing system for FAK
]
Wu, J. et al. (2015)
Decreased RECQL5 correlated with disease progression of osteosarcoma
. Biochemical and Biophysical Research Communications, Volume 467, Issue 4, 27 November 2015, Pages 617–622. [
AAVS1 safe harbor gene knock-in kit
]
Foss, O. et al. (2015)
Role of FOXA genes in maintaining transformed phenotype in human bronchial epithelial cells
. NTNU Open, 2015. [
FOXA1 gene, sgRNA, donor, AAVS1 safe harbor gene knock-in kit
].
Hu, B., et al. (2016).
A Novel Missense Mutation of CMT2P Alters Transcription Machinery
. Annals of Neurobiology doi: 10.1002/ana.24776
[LRSAM1 CRISPR sgRNA clone]
供体克隆服务
供体克隆设计与构建
我们会依据您的实验需求,提供供体克隆客户定制服务。供体克隆能将您感兴趣的基因、筛选标记或遗传因子通过CRISPR-Cas9介导的同源重组整合到宿主细胞的基因组内。我们提供一系列含有不同筛选标记及遗传因子的供体载体设计以满足您的实验需求。.
稳转细胞株服务
单克隆细胞株
含有CRISPR-Cas9介导的基因组修饰的单克隆细胞系。
为含有CRISPR-Cas9介导的基因组修饰的单克隆细胞系建库。
供体载体类型
LoxP 位点
pDonor-D01
copGFP
Puromycin
pDonor-D02
copGFP
Neomycin
pDonor-D03
Neomycin
pDonor-D04
Puromycin
pDonor-D05
Neomycin
pDonor-D07
copGFP
Puromycin/TK
pDonor-D08
copGFP
Neomycin/TK
pDonor-D09
Puromycin/TK
pDonor-D10
Neomycin/TK
pDonor-D11
copGFP
Puromycin/TK
pDonor-D12
copGFP
Hygromycin/TK
pDonor-D13
copGFP
Neomycin/TK
pDonor-D14
Puromycin/TK
IndelCheck™ CRISPR/TALEN 插入缺失检测体系及染色体水平功能验证服务
– GeneCopoeia 开发的 IndelCheck™ CRISPR/TALEN 插入缺失检测体系能用于 CRISPR/TALEN 的染色体水平功能验证及基因敲除细胞系筛选。我们同时也提供提供 CRISPR/TALEN 染色体水平功能验证服务。
Genome-TALER™ TALEN和TALE-TF定制服务
– GeneCopoeia提供多层次的TALENs,TALE-TFs和其他基于TALE的靶向基因组修饰工具的设计,构建和验证服务。
供体克隆定制及载体服务
– GeneCopoeia提供标准或客户定制的供体克隆载体,用于基因敲除、突变修饰、融合标签及其他方面的应用。我们提供带有不同元件的多种载体骨架,可供客户根据自己的实验需要进行选择。
基因组编辑稳定细胞系构建服务
– GeneCopoeia提供使用TALEN或CRISPR-Cas9技术构建的基因组编辑稳定细胞系。我们的稳定细胞系服务也可以兼容我们的人类及小鼠safe harbor基因敲入体系。
转基因小鼠服务
– GeneCopeia 提供原核显微注射法生成的带有 CRISPR 或 TALEN 介导的基因组修饰小鼠。
Genome-TALER™ 及 GeneHero™ 人类 AAVS1 Safe Harbor 基因敲入试剂盒及敲入克隆
——试剂盒能通过 TALEN 或 CRISPR-Cas9 介导,将您的目的基因、筛选标记及其他遗传因子特异敲入人类第19号染色体上
已发表文章
Schuler, B., et al. (2023).
Host Lipid Transport Protein ORP1 Is Necessary for Coxiella burnetii Growth and Vacuole Expansion in Macrophages.
mSphere doi: 10.1128/msphere.00104-23 [
GeneHero™ sgRNA expression clones targeting OSBPL1A
]
Pal, D., et al. (2023).
H4K16ac activates the transcription of transposable elements and contributes to their cis-regulatory function.
Nat Struct Mol Biol doi: 10.1038/s41594-023-01016-5 [
GeneHero™ human AAVS1 safe harbor gene knockin kit
]
Naoki, I-T., et al. (2023).
The Src-Family Kinases SRC and BLK Contribute to the CLDN6-Adhesion Signaling.
Cells doi: 10.3390/cells12131696 [
GeneHero™ sgRNA clones targeting mouse Blk; GeneHero™ sgRNA clones targeting mouse Scr
]
Talic, E-S., et al. (2023).
RNA Methyltransferase METTL16’s Protein Domains Have Differential Functional Effects on Cell Processes.
Curr Issues Mol Biol doi: 10.3390/cimb45070346 [
GeneHero™ sgRNA control; GeneHero™ METTL16 Cas9-expressing clone
]
Wan, Y., et al. (2023).
Nonmonotone invasion landscape by noise-aware control of metastasis activator levels.
Nat Chem Biol doi: 10.1038/s41589-023-01344-z [
GeneHero™ human AAVS1 safe harbor gene knock-in kits and clones
]
Schwarz, A.M., et al. (2023).
Terpenes from Cannabis sativa Induce Antinociception in Mouse Chronic Neuropathic Pain via Activation of Spinal Cord Adenosine A2A Receptors.
bioRxiv doi: 10.1101/2023.03.28.534594 [
GeneHero™ Cas9-expressing clone
]
Li, C., et al. (2023).
SUMO Proteomics Analyses Identify Protein Inhibitor of Activated STAT-Mediated Regulatory Networks Involved in Cell Cycle and Cell Proliferation.
J Proteome Res doi: 10.1021/acs.jproteome.2c00557 [
GeneHero™ PIAS1、PIAS2、PIAS3、PIAS4 sgRNA clones and negative control
]
Qin, Q., et al. (2023).
CNTNAP4 signaling regulates osteosarcoma disease progression.
NPJ Precis Oncol doi: 10.1038/s41698-022-00344-x [
T7 Endonuclease I Assay Kit; SuperCut™ Nuclease
]
Lin, P., et al. (2023).
TRAF6 regulates the abundance of RIPK1 and inhibits the RIPK1/RIPK3/MLKL necroptosis signaling pathway and affects the progression of colorectal cancer.
Cell Death Dis doi: 10.1038/s41419-022-05524-y [
GeneHero™ Cas9-expressing clone
]
Atobatele, A.G., et al. (2023).
Canonical and truncated transglutaminase-2 regulate mucin-1 expression and androgen independency in prostate cancer cell lines.
Cell Death Dis doi: 10.1038/s41419-023-05818-9 [
GeneHero™ TGM2 sgRNA clones
]
地址:广州科学城揽月路3号F区F801(510663)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}