3月24日,国际学术期刊《细胞研究》(Cell Research)在线发表了中国科学院上海生命科学研究院生物化学与细胞生物学研究所研究员许琛琦研究组与南京大学教授曹毅合作完成的最新研究成果Lipid-dependent conformational dynamics underlie the functional versatility of T-cell receptor。该研究揭示了TCR复合物胞内段的结构动态性,为TCR复合物传导不同刺激信号提供了结构基础。
在获得性免疫系统中,T细胞能够清除病原体和感染病变的细胞。T细胞抗原受体(T-cell receptor, TCR)是T细胞识别“自我”和“非我”物质的主要受体,可以和抗原呈递细胞(APC)表面特定的peptide-MHC复合物(pMHC)结合来发挥作用。ab TCR复合物由TCRab异源二聚体、CD3eg、CD3ed和CD3zz 组成。TCR与pMHC结合后,会引起TCR复合物中CD3分子胞内段免疫酪氨酸基序(immunoreceptor tyrosine-based activation motifs, ITAMs)中酪氨酸残基的磷酸化,随后开启下游信号。这个过程可以分为两个阶段,第一阶段是TCR由结构关闭到结构打开的转换,第二阶段是由TCR结构打开到ITAMs磷酸化的激活态。许琛琦之前的工作表明,CD3e和CD3z的胞内段含有碱性氨基酸富集区(Basic Rich Sequences, BRS,带正电),这些BRS可以通过电荷相互作用与富含酸性磷脂(带负电)的细胞膜内侧相互作用,使得其ITAM被磷脂保护起来,免于被下游的激酶磷酸化 (Xu et al., Cell, 2008)。随后他们发现,在T细胞活化初期,内流的Ca2+可以通过中和细胞膜内侧的酸性磷脂所带的负电荷,帮助那些未结合抗原的TCR复合物的胞内段从细胞膜上解离下来,从而发生磷酸化并放大初始的TCR信号 (Shi et al., Nature, 2013)。这些研究证明了细胞膜磷脂对TCR信号转导的重要调控作用。此外,TCR复合物在接受不同的抗原刺激时会产生不同的下游信号,从而引起不同的T细胞免疫反应,但产生这种功能多样性的结构基础依然不甚清楚。
为了探究该科学问题,在许琛琦、曹毅的联合指导下,博士后郭兴东、博士研究生闫成松、副研究员李华、博士研究生黄文茂等人运用多种技术手段,分析了TCR复合物胞内段与细胞膜磷脂结合的结构动态性。首先,他们利用单分子原子力显微镜(AFM),检测CD3e 胞内段(CD3eCD)与细胞膜内侧相互作用的力学特征。研究发现,CD3eCD从细胞膜解离的过程中会产生特异的力谱;除了单峰力事件外,还有一定比例的双峰力事件,这表明CD3eCD中很可能存在两个与细胞膜结合的位点。研究人员通过计算发现,除了之前鉴定的BRS区域外,在脯氨酸富集区 (Proline Rich Sequences, PRS) 和ITAM前半部分区域还存在一个较弱的次级膜结合位点。为了验证该推测,他们随后使用核磁共振(NMR),结合溶液PRE(solvent paramagnetic resonance enhancement)试剂TEMPOL,测定与磷脂结合的CD3eCD的构象动态特征。结果显示次级膜结合位点区域的氨基酸残基PRE效应相对较低,这进一步验证了次级膜结合位点的存在。而将该结合位点中的碱性残基和疏水残基突变后,发现CD3eCD前半段依旧与磷脂结合而后半段却从磷脂上解离下来。最后,他们利用全内反射荧光显微镜(TIRFM)和荧光共振能量转移(FRET)技术,检测不同强度抗原刺激下CD3eCD从细胞膜上解离的程度,发现在不同强度的抗原刺激下,CD3eCD从细胞膜上的解离程度的确存在差异。这表明,不同强度的抗原确实可以使CD3eCD处于不同的构象态。这些实验证明,膜脂依赖的TCR复合物构象动态特征很可能是TCR复合物传导不同刺激信号的结构基础。
该项研究得到了清华大学教授刘万里,上海生科院生化与细胞所研究员黄超兰、欧阳波,中科院武汉物理与数学研究所研究员唐淳,中科院长春应用化学研究所研究员王宏达,美国Lehigh University教授张晓辉的大力帮助,并得到国家自然科学基金委、科技部、中科院先导专项及上海市科委的经费支持。该研究工作还得到国家蛋白质科学设施(上海)核磁共振系统、质谱系统、复合激光显微镜系统、分子影像系统,生化与细胞所细胞分析技术平台、分子生物学技术平台、动物实验技术平台的大力支持。
文章链接
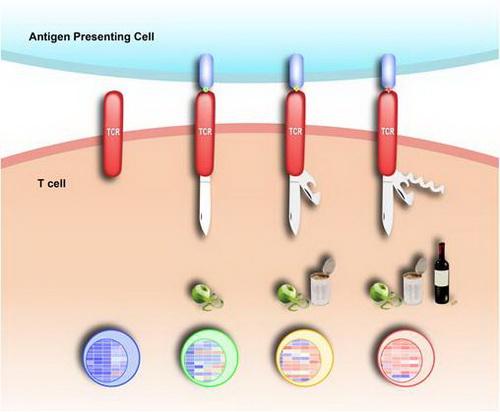
强弱不同的特异性抗原能够诱导TCR复合物产生不同的构象变化,从而产生不同的活化信号以及不同的免疫反应


