

「读完了文章的每个字,但不明白整段话的意思」,那我的大脑干了什么?
关注者
8,177
被浏览
282,493
164 个回答

谢
刘柯邀。


<Sir Joshua Reynolds, Dr Samuel Johnson, 1772, oil on canvas, 75.6 x 62.6 cm, Tate Britain, London.>
这个问题看似简单,实际是一个很Fundamental的问题,目前有的其他答案或多或少解答了一些,但是并不完整,合并起来也不足够解释问题。为了从根本上系统性的解释问题,我从1904年起的日俄战争讲起。


<Mosin–Nagant Model 91>
当时俄军使用的是莫辛-纳甘步枪(Винтовка Мосина, Mosin–Nagant Model 91),该枪使用7.62毫米步枪弹,弹头初速大概620m/s,穿透力很强,但是停止作用较弱。弹头射入人体之后不会造成太大的附带损伤,而是直接穿透。因此很多中弹的日军都能幸存下来,即使是头部中弹。
刚从东京大学毕业的眼科专家Tatsuji Inouye (井上達也)负责检查士兵的视觉损伤,以据此发放适当的抚恤金。

<Tatsuji Inouye (井上達也)>
但是井上達也并不满足于此,他注意到脑部的损伤部位跟视野的缺失部位有一定的对应关系,于是着手研究。为了客观的评估视野损伤的区域,他制作了颅脑坐标参照器(Kraniokoordinometer, cranio-coordinometer),以排除脑子大小对结果的影响,并准确评估视野区域。
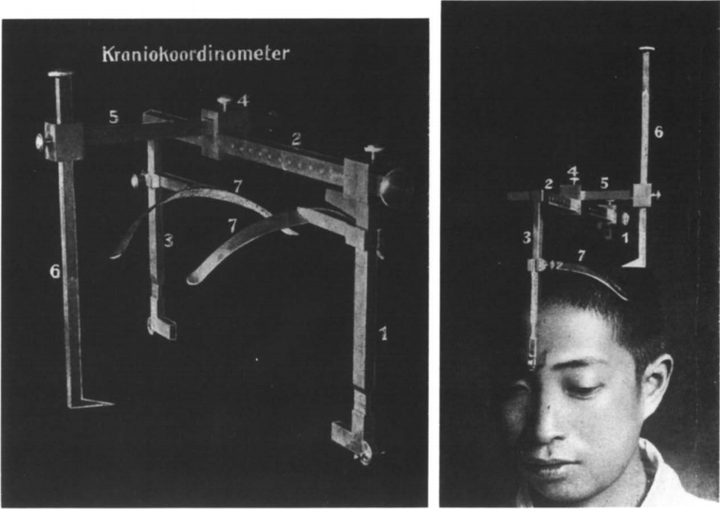

<颅脑坐标参照器,cranio-coordinometer>
他检查了28位典型头部中弹视觉损伤病患,测试的结果证实了他的想法——即脑部区域与视野存在着一定的对应关系[1],得到了世界上第一张Visual Cortex Map[2]
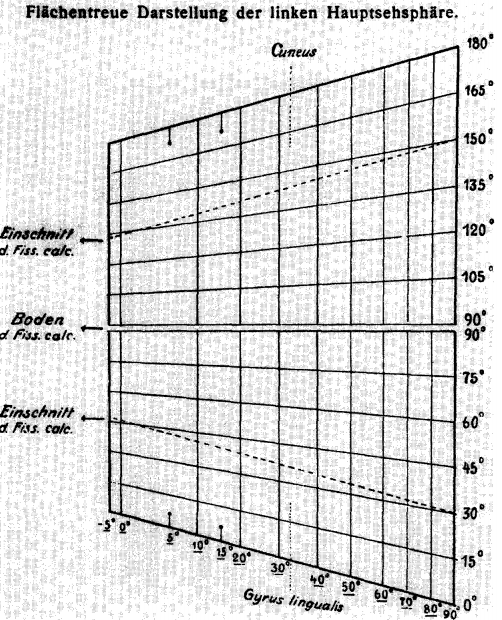

实验的结果以德文发表(Visual Disturbances following Gunshot Wounds of the Cortical Visual Area)[1,3,4]

论文发表后五年,第一次世界大战爆发了。这时候战争双方所使用的步枪武器特性跟日俄战争中俄军事用的类似,同盟国的德国使用的主要是Gewehr 1888(1888式委员会步枪)。


<Gewehr 1888>
当时在Royal Army Medical Corps(皇家陆军医疗队)服役(因为弱视的关系,他不被允许上前线,只允许在辅助部队服役)的神经解剖学家(Neuroanatomist) 英国神经解剖学家Sir Gordon Holmes(高顿-福尔摩斯 爵士)受到Tatsuji Inouye工作的影响,开始研究脑损伤(枪伤)跟视野损失的关系[5]。


<Sir Gordon Holmes, 左起第二,1915年>
在检查了2000多名伤兵之后,他着重对23名枕叶(后脑)受伤者做了分析。得益于X光机的实用化,Sir Gordon Holmes可以比Dr. Tatsuji Inouye更为准确的观察脑组织损伤的位置,从而绘制出更加完备准确的Visual Cortex Map, 也就是后来被称作Retinotopy(视网膜拓扑映射)的技术[6]. 此外,他还研究了脑外伤跟偏盲/象限偏盲的关系[7].
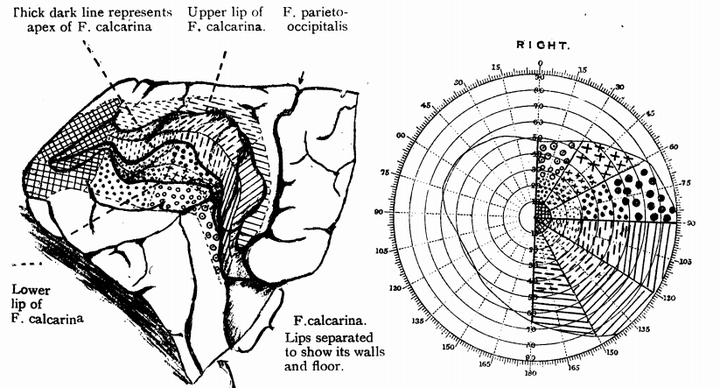

以上便是视觉皮层发现的基本过程[8]。视觉皮层也是神经解剖方面发现的第一个特定功能脑区,在这之后,感觉运动皮层,听觉皮层等相继被发现,均在很大程度上得益于战争中的脑外伤研究。
与此同时,在研究神经解剖方面,还有另外一派,他们通过研究脑皮层的细胞组织和髓鞘结构(Myelocytoarchitecture)来划归脑区,而不是根据脑区的功能。

<Cytoarchitecture: 6个月婴儿的大脑皮层的典型六层结构;引自论著[11]>
这一脉的重要创立者便是Prof. Oskar Vogt [9].

<Prof. Oskar Vogt>
他于1914年创立了威廉皇帝学会脑科学研究所(Kaiser-Wilhelm-Institut für Hirnforschung), 也就是先今的马普学会脑科学研究所(Max Planck Institute for Brain Research, Max-Planck-Institut für Hirnforschung). Oskar Vogt曾在此研究过列宁的脑子。1959年,他又与他的妻子Cécile Vogt-Mugnier创立了Cécile and Oskar Vogt Institute of Brain Research (又被称作Vogt-Vogt school). 该研究所后被杜塞尔多夫大学(
C辿cile and Oskar Vogt Institute of Brain Research)吸收,是目前大热的Big Brain Project [10]初创方(
Forschungszentrum J端lich),这是后话了。
Oskar Vogt的高徒Korbinian Brodmann(布罗德曼),从1901年开始跟随他工作。


<左一: Korbinian Brodmann; 左二: Cécile Vogt-Mugnier, 左三/中间大胡子者: Oskar Vogt >
Brodmann在Oskar Vogt的指导下,花费数年观察大脑皮层的解剖结构(Cytoarchitecture and Myeloarchitecture), 在1909年发表了他的大作"Vergleichende Lokalisationslehre der Großhirnrinde in ihren Prinzipien dargestellt auf Grund des Zellenbaues" (Comparative Localization Studies in the Brain Cortex, its Fundamentals Represented on the Basis of its Cellular Architecture)[11].
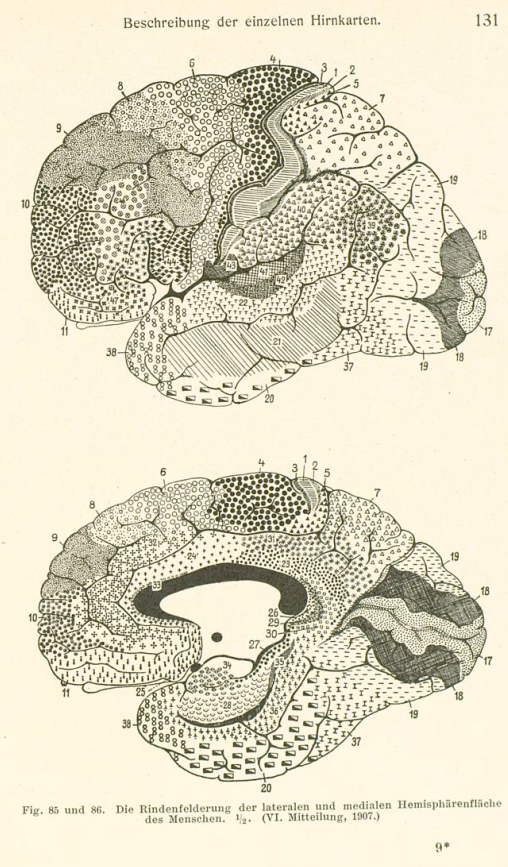

<原著中的Brodmann分区[11]>
根据细胞组成结构将大脑皮层划分为52的区域,其中关于枕叶皮层(视觉)的划分跟Sir Gordon Holmes的结果具有相当的吻合性,可谓殊途同归。
Vogt-Brodmann这套方法可以称为 结构 派[12],Inouye-Holmes方法可称为 功能 派。这两种方法随着技术(EEG, CT, MRI, MEG, TMS,PET,PLI,Immunohistochemistry)的发展,都逐渐演化精进,构成我们今天脑科学/神经科学的枝干。这两派的发起,可以称之为神经科学/脑科学的 一次革命 ,我们对大脑逐渐从一无所知到略有小成,伴随着这个进程的,是科学研究的中心从德国转移到美国,科学的语言也从德文转变为英文。我们对视觉通路的认识,不再局限于视野和枕叶的遥远关系,而是直接的联系起来[13]:
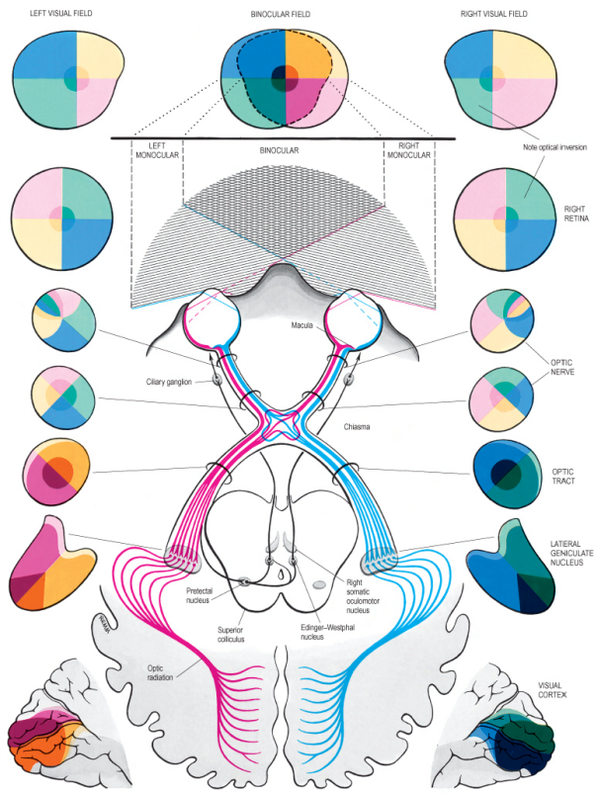

<视觉通路:视野与视觉皮层的联系>
人脑的视觉信号传导通路,在进入大脑皮层之前是高度的结构化的、分工明确的。视网膜感知的物体,通过该通路,会以一定空间关系映射视皮层上[14], 该关系也就是上文中提到的视网膜拓扑映射(Retinotopy)[15].
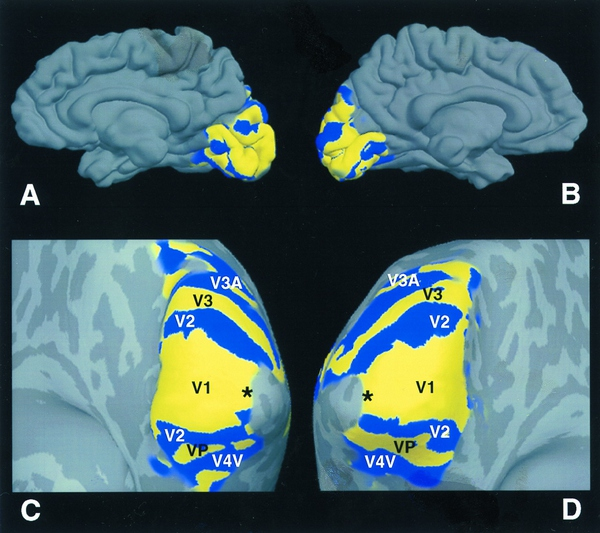

<Retinotopy>
信息在进入初级视觉皮层之后,视觉信号的处理会会表现出非常复杂的状态。根据结构和功能的信息,以及MRI技术帮助,发现视觉信号传递到初级视皮层V1之后,会继续向更高级的视觉皮层传递,据此可以将视觉皮层更精细的分类,同时根据信息传递的关系,得到它们内部之间的分级结构。
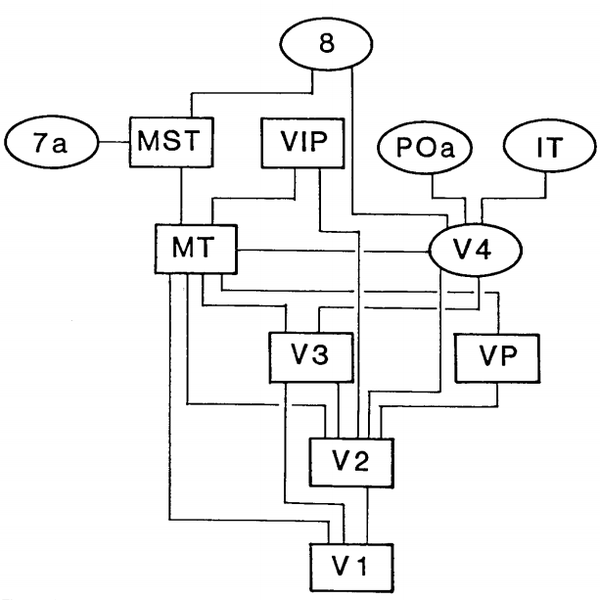

<视觉皮层分级结构>
从初级到更高级的视觉皮层,视觉信息逐级传递。人脑理解的内容越来越复杂化、抽象化,由“模式”变成具体的"物",再到物的特性和物与物之间的关系。
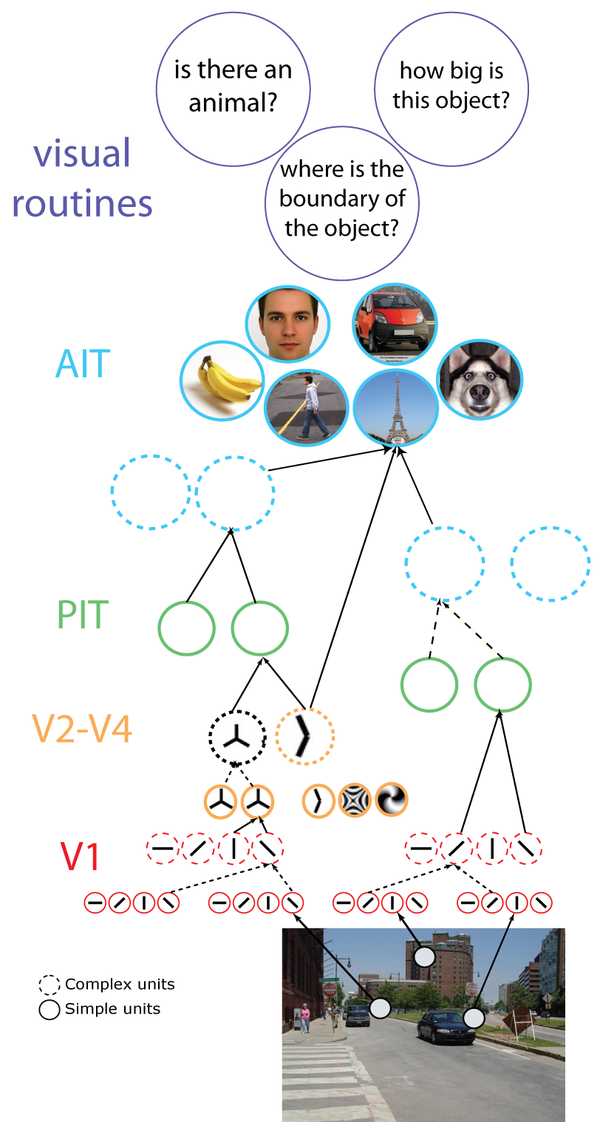

<视觉分级结构的功能分工>
在逐级传到过程中,人们也注意到,其在皮层的传导可以大体分成两个通路,腹侧通路(Ventral Pathway/Stream)和背侧通路(Dorsal Pathway/Stream)[17]。
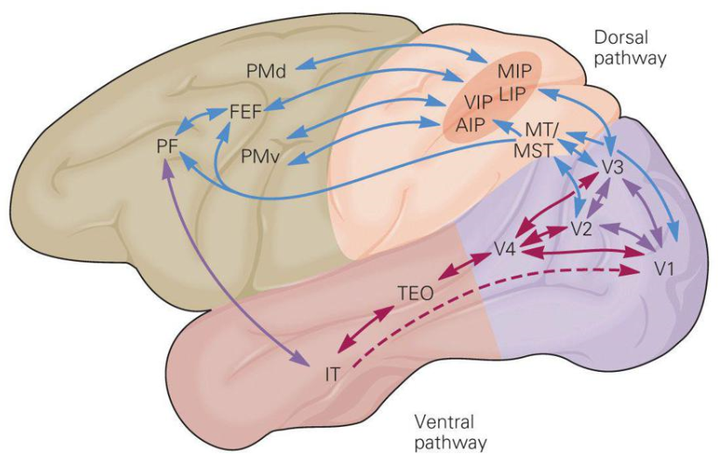

分别又按照功能,被称作“What”和“Where”通路:

其中“Where”被认为跟物体的位置和运动处理有关,“What”被认为跟物体的识别有关[18]。当然,这种说法/猜想缺乏足够的证据支持,仍然饱受争议。在上面的视觉信息传递图和分级结构图中,我们可以看到, 信息在进入人脑初级视觉皮层之后,其传递的过程非常的复杂,涉及到几乎全脑的区域之间的写作。语言区域的引入自然是不可少的,而当我们在读的相应的内容,比如关于运动的内容,就需要有运动相关脑区的引入;对应的,读到情感内容,就需要有情感相关脑区的工作协同。我们在想象一件事情时,大脑中的工作模式,跟我们实际操作这件事情时非常类似 。
神经科学发展到这里,已经可以部分的回答这个问题,即所谓的" 视而不见 "," 读而不解 ",是 我们的大脑在工作之中缺少必要的协同,相关的脑区并没有进入工作状态 。类似的,我们在学习某一项技能/学科之前,由于脑内还没有构建好协同工作状态,以及特定脑区的功能不足,我们在读这些相关图书内容的时候,也是会读而不解,即使我们想尽办法集中 注意力 也无济于事。以上也是目前的心理学/神经科学领域教科书能给出的最新的回答。
但是,这个问题还是没有得到足够的回答,如要问什么是特定的工作协同状态,使用以上的知识/事实就显得不够可靠了。当然神经科学到此并没有停止发展,而是发生了一定的质变。
以上相当部分的认识,在MRI/fMRI引入神经科学研究之后发现的。随着研究的深入,神经科学家不可避免的要碰到两个问题:
- fMRI的信噪比低,即使使用超高场设备还是显得不足
- 人脑的个体差异极大,人与人之间,甚至同一个人的左脑和右脑之间的差异有时候都不可忽略。
以上两个问题中,1可以通过技术进步得到改观,而2则是目前fMRI研究不可避免的致命缺陷,这就造成相当部分的fMRI研究的结果重复困难,可以说目前大部分已经发表的fMRI研究结果都是不可信的,尤其是心理学方面的fMRI研究,由于涉及更为高级神经功能,其可靠性更是存疑,为了解释奇怪的结果,一些稀奇古怪的说法也能编出来。fMRI研究要得到想要的或者是好的结果,一方面实验设计不能出问题,另一方面 一定的运气也是不可缺少的 。那些只靠一个fMRI实验就做出结果顺利毕业的博士研究生同学,必然是祖上积了八辈子德了。
越是高级的功能区,个体差异越大(位置和大小, 甚至有无),近代神经科学从视觉皮层的研究开始并非偶然,视觉皮层初级功能区是我们人类大脑中最大的一块功能相对单一的特定功能区,这一区域面积/体积大,个体差异相对小,区域的结构和功能边界也相对明显并且比较一致。其他相对小一些的功能区,比如感觉运动皮层区域、初级听觉皮层区域,也有比较好的个体间一致性。一旦牵涉到所谓高级功能区域,比如视觉的V5及更高区域,上面Where和What通路的大部分区域,个体间的差异对统计结果的影响因素就不可避免了。但是行为实验,病理实验都需要做组间对照和统计,所谓区域与区域间的比较,很可能就用错了位置,致使组间统计的基础都不够牢固,引入更多被试可能平均掉一些个体差异,但也可能是引入了更多变量,致使差异不收敛。其结论无论是阴性还是阳性,都要大大的打个问号。
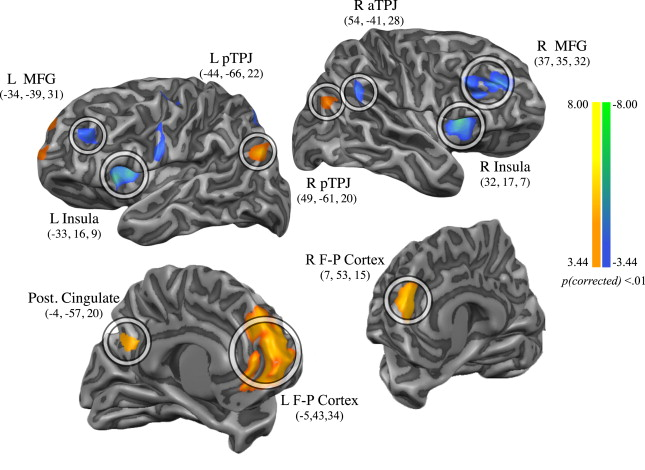
为了解决或者回避个体差异的根本问题,我们必须否定前面的认识,即关于个体脑组织结构的基本假设,也就是说,Vogt-Brodmann基于细胞组织结构的分区,Inouye-Holmes基于功能的分区,也是需要重新商榷的。 脑部各区域之间的界线,并不是明显的,而是模糊的;并不是静止的,而是可变的;并不是相互独立的,而是可以互相包含,犬牙交错的 。同时,脑部区域的功能,也不是一层不变的 。以上这些特征,使得目前的统计方法在很多方面并不适用于神经科学研究。我们在 思考神经科学问题时,与其考虑脑区,不如考虑功能/结构节点 ,这也就是目前神经科学领域正在发生的 二次革命:Connectome(脑连接体) .
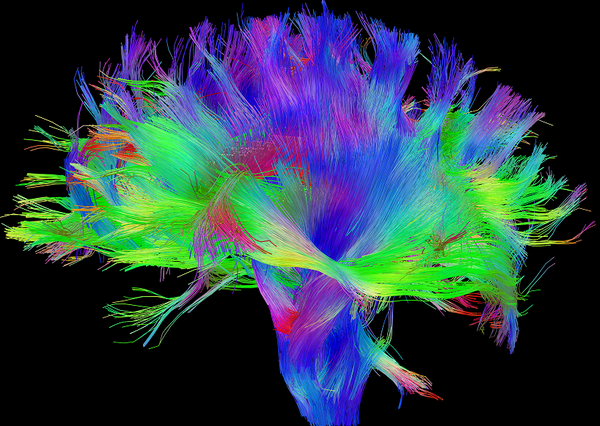

人脑在结构上存在一个人眼 可直接观察的连接体:白质纤维束连接 。这一连接体把整个大脑通过神经元的轴突和树突连接起来。这一结构虽然同样的存在很大的个体间差异,但是因其是物理性的连接,在短时间内存在一定的稳定性,这是我们大脑连接体的硬件。另外还存在一个人眼 不可直接观察的连接体:功能连接 ,即我们大脑区域间的协同工作机制,是我们大脑连接体的软件。
在我们的大脑内,在神经元的突触前膜向突触后膜起信息传递作用的化学物质。按照作用后果可分为离子型(Ionotropic)和代谢型(Metabotropic)两类。其中离子型受体按照电位变化可分为 兴奋型 和 抑制型 两类。这些神经递质的分泌情况,构成了大脑区域间功能连接的柔性导线,它们不断的联通(强化性连接)和切断(抑制性链接)连接的开关,形成大脑在应对不同任务时候的各自工作模式。
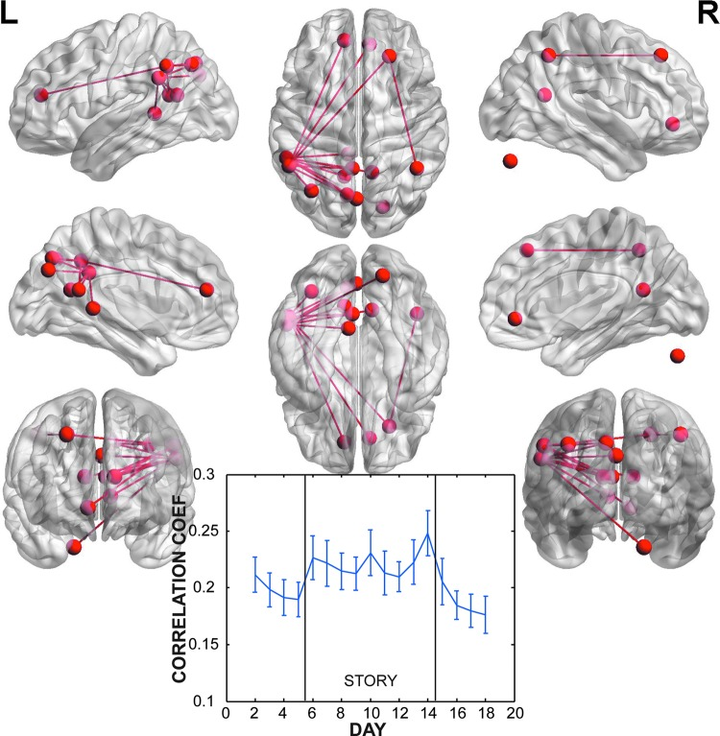

通过测度阅读任务者,我们可以得到该任务造成的脑内连接的改变情况, 这种连接情况的改变,可以解释为阅读能力的改变,其实是我们大脑进行阅读任务时功能连接的残影 [19]。
进行高级任务时,控制功能连接柔性导线的系统中,一个重要的因素是我们意识的注意力(attention)[20]。

<在高级任务中,注意力对功能连接的主动控制>
到这里,就可以更为准确的解释楼主的问题了,所谓读不懂,原因有二:
- 注意力没有集中,脑中任务处理的连接网络没有足够的激活水平
- 缺乏相关的知识/背景/经验,脑中任务处理机制网络的能力没有成熟
目前神经科学领域的这第二次革命才刚刚开始,很多研究在依赖第一次革命的既定成果的同时,还受到其局限性的限制,比如在连接节点上,还经常性的受限于以往固定性的脑区划分,而不是按照个体内脑区功能做划分。当然,如果按照后者,在统计上就存在既定的困难。在其他领域,比如人工智能领域,我们对人脑的模拟最多还停留在使用第一次革命结果的阶段,我们目前使用的智能方法,包括神经网络、机器学习/深度学习,都是把处理机制相对的固定化,因而其方法只对特定的任务/特定的数据好,自应能力非常苍白。目前对人工智能取代人脑的担忧,只是杞人忧天罢了。
以上。
--------
[1] Inouye, T. "Die Sehstoerungen der kortlichen Sehsphaere. W." (1909): b8.
[2] Glickstein, Mitchell, and David Whitteridge. "Tatsuji Inouye and the mapping of the visual fields on the human cerebral cortex." Trends in Neurosciences 10.9 (1987): 350-353.
[3] Tubbs, R. Shane, et al. "Tatsuji Inouye: the mind’s eye." Child's Nervous System 28.1 (2012): 147-150.
[4] Adams D, Horton J (1991) Book reviews. Nature 412:482–483
[5] McDonald, Ian. "Gordon Holmes lecture: Gordon Holmes and the neurological heritage." Brain 130.1 (2007): 288-298.
[6] Holmes, Gordon. "Disturbances of vision by cerebral lesions." The British journal of ophthalmology 2.7 (1918): 353.
[7] Holmes, Gordon. "Disturbances of visual orientation." The British journal of ophthalmology 2.9 (1918): 449.
[8] Glickstein, Mitchell. "The discovery of the visual cortex." Scientific American 259.3 (1988): 84-91.
[9] Vogt, Cécile, and Oskar Vogt. Allgemeine ergebnisse unserer hirnforschung . Vol. 21. JA Barth, 1919.
[10] Amunts, Katrin, et al. "BigBrain: an ultrahigh-resolution 3D human brain model." Science 340.6139 (2013): 1472-1475.
[11] Brodmann, Korbinian. Vergleichende Lokalisationslehre der Grosshirnrinde in ihren Prinzipien dargestellt auf Grund des Zellenbaues . Barth, 1909.
[12] Brodmann, Korbinian. "Neuere Forschungsergebnisse der Großhirnrindenanatomie mit besonderer Berücksichtigung anthropologischer Fragen." Naturwissenschaften 1.46 (1913): 1120-1122.
[13] Standring, Susan. "Gray’s anatomy." The anatomical basis of clinical practice 39 (2008).
[14] Tootell R B H, Hadjikhani N K, Vanduffel W, et al. Functional analysis of primary visual cortex (V1) in humans[J]. Proceedings of the National Academy of Sciences, 1998, 95(3): 811-817.
[15] Engel S A, Glover G H, Wandell B A. Retinotopic organization in human visual cortex and the spatial precision of functional MRI[J]. Cerebral cortex, 1997, 7(2): 181-192.
[16] Van Essen, David C., and John HR Maunsell. "Hierarchical organization and functional streams in the visual cortex." Trends in neurosciences 6 (1983): 370-375.
[17] Kandel, Eric R., James H. Schwartz, and Thomas M. Jessell, eds. Principles of neural science . Vol. 4. New York: McGraw-Hill, 2000.
[18] Ungerleider, Leslie G., and James V. Haxby. "‘What’and ‘where’in the human brain." Current opinion in neurobiology 4.2 (1994): 157-165.
[19] Berns, Gregory S., et al. "Short-and long-term effects of a novel on connectivity in the brain." Brain connectivity 3.6 (2013): 590-600.
[20] Tyler, Sarah C., et al. "Functional connectivity of parietal cortex during temporal selective attention." Cortex 65 (2015): 195-207.

7月2日更新
更新为两件事:一,推荐一下
@MIAO TONG从神经方面的回答。二,评论中有些对我给出的例子的误解,稍作解释。
一,目前赞同数最高的答案作为对神经科学发展的科普很棒,但对本题来说不太合适。MIAO TONG的答案并没有像其开篇说的那样不完整,我也并不认为高票答案“从根本上系统性”地解释了问题。以下提出几个点:
(1)先看这个高票答案最后给出的结论,说“读不懂”有以下两个原因:
- 注意力没有集中,脑中任务处理的连接网络没有足够的激活水平
- 缺乏相关的知识/背景/经验,脑中任务处理机制网络的能力没有成熟
这两点总结得很对,但它们不是直接从答案中直接develop来的,至少只占了很少的篇幅。首先这个答案中对注意的作用的描述只有两句:
进行高级任务时,控制功能连接柔性导线的系统中,一个重要的因素是我们意识的注意力(attention)
<在高级任务中,注意力对功能连接的主动控制>
注意会影响高级任务的表现这一点是个众所周知的事实,注意不足时各脑区/功能间连接不够“有效”是很自然的,但注意具体怎样影响阅读这个任务答案里没有说。但MIAO TONG的回答里最后一段说明了:
另一种阅读方法是更耗能地集中注意力,积极发动高级思维结构,由字到词汇、词组再到语句进行信息的理解和存储,遇到不懂的地方果断去查资料。如果上述两种方法都不采用,不反复使用语音辅助,也不集中注意力去思考,那么就永远无法理解了。
(2)高票答案说读不懂是相关网络的能力没成熟没有问题,但前半句“缺乏相关的知识/背景/经验”与后半句“脑中任务处理机制网络的能力没有成熟”之间的因果关系在回答中是如何发展而来的?我确实没找到。坦白说,我觉得这个缺乏概念关系说明、直接给出结论的点是这个回答作为“有效答案”的致命伤。顺带一提,之所以要反复读复杂的文章,一个重要原因就是集中注意。
(3)另外对于“网络”MIAO TONG的回答里也有:
题主的情况 是晦涩难懂的语音信息在感觉通路中的短暂停留,形成的记忆是瞬时记忆,可能角回与Wernicke’s 区也进行了初步的分析,然而海马、额叶等高级思维结构却没有很好地参与其中。
所以我认为说她的答案不完整并不公平,她只是没把这两点总结出来。
(4)首先用大篇幅讲了脑功能定位论(脑区划分等)的发展、视觉通路的功能(知觉方面很重要,但不是语言方向研究的重点),然后从fMRI等新兴技术的角度阐述了定位论的不足,进而引入对脑连接体的描述——这是我对高票答案的概括。但它关于“读不懂”的神经机制的解释真的不多,如前所述,结论很好但不是从答案中的描述直接发展而来的。
未免有挑衅之嫌就不@该答案的答主了,另外这个答案作为神经科学发展的科普还是挺好的,放在这儿被大家看到也是物尽其用。
二,我主要是从“缺少相关知识背景导致读不懂”这个角度回答的,所以举了下文中的“ABCD”例子。评论中有同学对这个例子提出质疑,大意是觉得我给出的A、B、C、D可以有多种指代,所以本身就推理不出它们究竟是什么,于是觉得这例子不恰当。
但我想说的就是这个意思啊……在缺少相关知识等情况下,很难将作者文中的概念与自己对事物的已有知识联系起来,于是难以提取不同概念间的逻辑关系。正因为不知道ABCD是什么,才觉得它们“什么都可以是,又什么都不是”,所以才读不懂啊。
--------------------------------------------------------------------------------------------------------------------------------------------
题主描述的很清楚,这种反复读才能明白甚至最终也不明白的事一般会发生在读“生涩论文”的时候。此时读者对每个字、词的意思是清楚的,但是不能有效将它们联系起来,无法在自己的思维体系里整理出各概念间的关系。
这可能是因为读者对文章中的概念(或者说词汇在作者的思想体系中指代什么)不熟悉;也可能是因为不能清晰理解作者想阐述的概念与概念间的逻辑关系。
自然科学类文章就很容易出现概念的问题,我写的一些答案就经常被说读不懂,但在相关领域有知识基础的同学就不会有这种感觉。 概念指代不清楚就无法将其整合进自己已有的知识体系,也就无法提取出它们之间的关系。问题出现在“表征”或“联结”上。 我不是语言方向的,不知用这两个词合不合适,望指正。
比如我随便说两句话:3只C1与4只C2在A处吃B,不远的A‘处还有5只C1和7只C2在休息。与此同时一只D1与一只D2正看着它们。
然后问,(1)C1和C2有什么区别?(2)D1和D2打算干什么?
不知道有多少人愿意回答这种问题。但如果我换种方式说:
3只公牛与4只母牛在树旁吃草,不远的另一棵树旁还有5只公牛和7只母牛在休息。与此同时一只公狮和一只母狮正看着它们。
显然,(1)性别不同。(2)吃饭。
如果我不用“只”做量词,那这场面其实也可以变为老张一大家子在吃东西,与此同时老李两口子正看着他们。
好像有点儿扯远了,总之在概念不清的时候读到的东西就不是草原上两头狮子对着一群牛垂涎欲滴的画面,而是一堆不知道什么东西的ABCD,谁在吃谁在看转眼就忘了,更别提提取主旨回答问题了。
所以反复读的一个作用就是查明不清楚的概念,然后在不断重复的过程中对它更熟悉(在工作记忆中更好地储存等),那也就更容易在之后figure out概念间的关系了。
当然即使概念全都清楚也可能会读不明白,这可能就是因为概念间的关系网很复杂,但作者又无法简明地指出这些关系。有时这也不是作者的问题,毕竟概念多、关系复杂时这些东西就超出人类工作记忆的负荷了。另外很多哲学理论也会给人深奥的感觉,我不是个能把它们都读明白的人,也就不好分析读不懂的原因了。
至于在读不懂的情况下大脑干了什么:
(1)一些皮层下结构和初级视皮层要对接收到的视觉信号做最基本的加工,比如线条是横还是竖
(2)更高级的视皮层要识别语词,并将其与脑中的“词典”(visual input lexicon)相对应,知道每个词是哪个,比如自己不认识的词就找不到对应的
(3)内侧额叶要做语义方面的处理